Die Höhe der biologischen Fruchtbarkeit des Menschen ist nicht zufällig, sondern ein Ergebnis der biologischen Evolution und damit auf die natürlichen Verluste einer weitgehend von natürlichen Gegebenheiten bestimmten Umwelt unserer Vorfahren abgestimmt. Es ist anzunehmen, dass bei ihnen mindestens acht bis zehn Geburten notwendig waren, um die Art Mensch in der Auseinandersetzung mit der Umwelt zu erhalten (MOHRIG 1976, S. 53), womit auch von einer "Bestandserhaltungsfruchtbarkeit" gesprochen werden könnte. Sie schließt mit ein, dass unter den existentiellen Bedingungen während unserer biologischen Konstituierung nur zwei bis drei Kinder das geschlechtsreife Alter erreichten und zur Fortpflanzung beitragen konnten. Mit Beginn der sozialen Evolution erhöhte sich der generative Effekt einer jeden Geburt und hat bis in die Gegenwart der hochentwickelten Industrie- und postindustrialisierten Staaten ein solches Niveau erreicht, dass im Durchschnitt pro Frau "lediglich" 2,18 Geborene nötig sind, um den Bestand der entsprechenden Gesellschaften zu garantieren.
Damit unterscheidet sich die Bestandserhaltungsfruchtbarkeit von der biologischen Fruchtbarkeit seit Beginn der Sozialisation des Menschen und kann als soziale Fruchtbarkeit bezeichnet werden. Diese hat noch eine zweite Seite: Die durch den Menschen bewusst vorgenommene Regulierungen der Nachkommenszahl zur Erhöhung oder Verringerung der tatsächlichen Kinderzahl pro Frau.
Inwieweit biologische und soziale Momente der Fruchtbarkeit ineinander greifen, ist auch an der Sterblichkeit der für die Reproduktion einer Generation überzählig zu gebärenden Kinder zu erkennen. Diese ist keineswegs als eine "natürliche" Sterblichkeit in Form einer biologisch angelegten allgemeinen "Lebensuntüchtigkeit" anzunehmen, sondern Resultat vielfältiger biographischer Erscheinungen mit entsprechender Schwächung der Vitalität bis zur Todesfolge, wie z. B. epidemische Krankheiten und Hungerkatastrophen sowie selektive Auseinandersetzungen. Sie kommt in den generativen Relationen der von uns durch Dokumentation schon gut überschaubaren früh- bzw. vorindustriellen Entwicklung recht gut zum Ausdruck. So hatten die sozial schwächer gestellten Schichten der Bevölkerung in Oberbayern, wie durch Auswertung des Familienkatasters der Gemeinde Dietramzell gezeigt wurde, im Hungerjahr 1817 eine Kindersterblichkeit von fast 80 % (PHAYER 1970). Die etwa 20 % Überlebenden dürften entweder selbst in ihrem sozialen Umfeld noch privilegiert gewesen sein oder sie hatten einfach eine bessere biologische Kondition, obgleich beide Sachverhalte auch in wechselseitiger Abhängigkeit gewirkt haben dürften.
Der in Teilen der Demographie üblichen weiteren, fast hierarchischen Differenzierung der Fruchtbarkeit nach natürlicher ehelicher, ehelicher und tatsächlicher Fruchtbarkeit als kleinste Teilmenge (vgl. MÜNZ/ULRICH 1994, S. 12) kann aus bevölkerungsgeographischer Sicht nicht gefolgt werden. Allein die Kategorisierung von ehelicher Fruchtbarkeit als "natürlich" anstatt "in der Ehe üblich" sowie die erheblichen regionalen Unterschiede in der Bedeutung der Ehe für die Geburt von Kindern zwingen zu einer anderen Ordnung der Begriffe. Wir gehen davon aus, dass eheliche und außereheliche Fruchtbarkeit nur zueinander komplementäre Teilmengen der tatsächlichen Fruchtbarkeit sind, denn das Recht und die Möglichkeit zur bewussten Steigerung oder Senkung der Geburtenzahlen, genauer: das Recht auf Vollendung einer geplanten Schwangerschaft, sollten heute alle Frauen unabhängig von ihrem Familienstand besitzen. Insofern ist auch der in der offiziellen deutschen Statistik nach wie vor gebräuchliche Begriff der "Legitimität" von Kindern in Abhängigkeit vom Familienstand der Mutter kritisch zu hinterfragen.
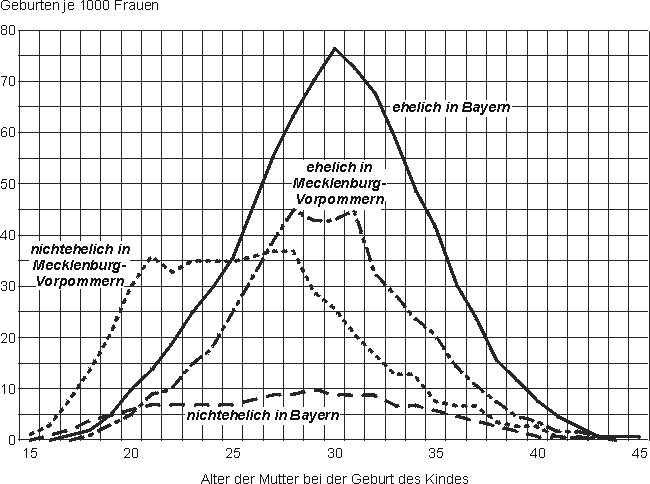
nach: GRÖNER 2000, S. 160